


Durch eine Fehlregulation in der Achse Hepcidin-Ferroportin lassen sich alle bekannten Formen der erblichen
Eisenspeicherkrankheit auf einfache Weise erklären. Bei verschiedenen Hämochromatoseformen liegt ein
Hepcidinmangel vor, sodass HFE, TfR2 und Hämojuvelin in der Leber direkt etwas mit der Hepcidinsynthese
zu tun haben müssen. Besonders Patienten mit HJV-Mutationen weisen extrem niedrige Hepcidinspiegel auf,
sodass HJV vermutlich der wichtigste Hepcidinmodulator ist.
Regulation der Hepcidinaktivierung
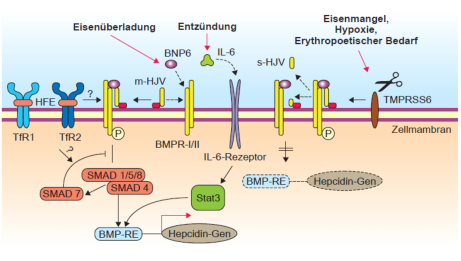
Hämojuvelin verursacht eine seltene, aber sehr schwere Form der erblichen Eisenspeicherkrankheit und ist
offenbar der wichtigste Aktivator der basalen Hepcidinaktivierung (67). HJV gibt es in einer membran-
assozierten Form (m-HJV) und einer löslichen Isoform (s-HJV). m-HJV ist ein Ko-Rezeptor für bone-
morphogenetic Proteine (BMPs), Zytokine, die die Hepcidin-Expression in vitro und in-vivo in Mäusen
stimulieren (68). m-HJV verstärkt das Signal, dass durch Bindung von BMP an den zugehörigen Rezeptor
generiert wird. Wahrscheinlich ist BMP6 der endogene Regulator der Hepcidin-Expression und wird
möglicherweise zukünftig eine wichtige diagnostische Rolle spielen (69). Das lösliche HJV ist eine
inhibitorische Komponente des Hepcidinweges. s-HJV wird bei Eisenmangel oder bei Hypoxie wahrscheinlich
von Muskelzellen freigesetzt.
Eine weitere neue Entdeckung ist TMPRSS6, das für eine TypII-Plasmamembran Serin-Protease, Matriptase-
2, kodiert. Matriptase-2 ist hoch konserviert im Mensch, Maus und Ratte und hoch exprimiert in der Leber.
Ein Mausmodell mit Eisenmangelanämie und Körperhaarausfall wurde von Ernest Beutler auf dem ASH
Meeting Ende 2007 vorgestellt, das eine Mutation im TMPRSS6 aufweist (70). Diese Mask-Mäuse können die
intestinale Eisenabsorption nicht hochregulieren und haben inadäquat hohe Hepcidinwerte, die die
Eisenabsorption blockieren und Ursache der Anämie sind. Momentan wird TMPRSS6 als Eisensensor bei
Eisenmangel angesehen, der das Signal der basalen Hepcidinaktivierung über BMP/BMP-R und SMAD
abschaltet. Ein mögliches Modell ist in Abb. 5 dargestellt.
Unklar ist weiterhin, über welche Mechanismen HFE und TfR2 auf die Hepcidinaktivierung einwirken. Die
Funktion der Matriptase-2 ist beim Menschen konserviert. Erste Studien berichten über einige Familien und
wenige sporadische Fälle mit eisenrefraktärer Eisenmangelanämie infolge TMPRSS6-Mutationen (71, 72).
Auf solche (seltenen) Fälle ist in der Abklärung von unklarer Eisenmangelanämie zukünftig zu achten.
Abb. 5 Hypothetischer Mechanismus der Funktion von TMPRSS6 in der Hepcidinaktivierung über HJV, BMP und
BMPR. Im Eisenmangel (Schere offen) wird die Hepcidinaktivierung abgeschaltet und es kann vermehrt Nahrungseisen
absobiert werden. Bei mutiertem TMPRSS6 (MASK-Maus) bleibt die Hepcinaktivieung bestehen und es kommt zu einer
Eisenmangelanämie durch ständige Hepcidinaktivierung. Bei Infekt/Endzündung wird über IL6 der IL6R aktiviert. Über den
Signalkaskadeweg STAT3 („signal transducer and activator of transcription 3”, Akute-Phase-Response-Factor) wird ein
regulatorisches Element im Hepcidinpromotor aktiviert (P.Nielsen Kap. 32 in Allgemeine und Spezielle Pharmakologie und
Toxikologie, Elsevier 2013).
Hepcidin bei Eisenmangel und Eisenüberladung
Die Grundzüge der Hepcidinaktivierung gelten heute als bekannt. Hepcidinmangel ist Ursache der erblichen
Eisenspeicherkrankheiten Typ1-3 und ist bei sekundärer Eisenüberladung theoretisch hochreguliert, sodass
kein Eisen mehr aus dem Darm aufgenommen wird (3). Bei Eisenmangel ist Hepcidin niedrig. Bei
Infekt/Entzündung ist bei Patienten mit Anämie ist die Hepcidinkonzentration im Serum meist hoch, was aber
nicht auf einzelnen Patienten zutrifft, bei denen offenbar noch andere Effekte wirken, die Hepcidinsynthese
oder – abbau beeinflussen (62). Insgesamt gibt es zum Thema Hepcidin inzwischen unzählige Studien bei
einzelnen Krankheitsbildern. Ein großes Problem war bisher die zuverlässige Bestimmung von Hepcidin im
Serum und im Urin. Die ist zum einen methodisch bedingt – es gibt keine guten Antikörper gegen das kleine
Peptid-, zum andern scheinen auch Tagesschwankungen und eine Tag-zu-Tag-Variation eine Rolle zu spielen.
Auch fehlen standardisierte Bedingungen der Probengewinnung und Verarbeitung. Es bleibt daher
abzuwarten, inwieweit Hepcidin den Weg in die Alltagsroutine der Diagnostik bei Eisenmangel und
Eisenüberladung finden wird.
Literatur Eisenstoffwechsel
1.
WHO/NHD/01.3. Iron deficiency anemia assessment, prevention, and control. World Health Organization, 2001
2.
Pietrangelo A. Hereditary hemochromatosis--a new look at an old disease. N Engl J Med 2004;350(23):2383-97
3.
Heilmeyer L, Plötner K. Eisenmangelzustände und ihre Behandlung. Klin Wochenschr 1936; 15:1669
4.
Feder JN, Gnirke A, Thomas W, Tsuchihashi Z, Ruddy DA, Basava A, Dormishian F, Domingo RJr, Ellis MC,
Fullan A, Hinton LM, Jones NL, Kimmel BE, Kronmal GS, Lauer P, Lee VK, Loeb DB, Mapa FA, McClelland E,
Meyer NC, Mintier GA, Moeller N, Moore T, Morikang E, Prass CE, Qiuntana L, Starnes SM, Schatzmann RC,
Brunke KJ, Drauna DT, Risch NJ, Bacon BR, Wolff RK. A novel MHC class I-like gene is mutated in patients
with hereditary haemochromatosis. Nature Genetics 1996; 13:399-408
5.
Bothwell TH, Charlton RW, Cook JD, Finch CA. Iron metabolism in man. Blackwell Scientific Publications, 1979
6.
Brock JH, Halliday JW, Pippard MJ, Powell LW. Iron metabolism in health and disease. WB Saunders
Company Ltd. London , Tokyo 1994
7.
McKie AT, Barrow D, Latunde-Dada GO, Rolfs A, Sager G, Mudaly E, Mudaly M, Richardson C, Barlow D,
Bomford A, Peters TJ, Raja KB, Shirali S, Hediger MA, Farzaneh F, Simpson RJ. An iron-regulated ferric
reductase associated with the absorption of dietary iron. Science 2001; 291:1755–1759
8.
Gunshin H, Mackenzie B, Berger UV, Gunshin Y, Romero MF, Boron WF, Nussberger S, Gollan JL, Hediger M.
Cloning and charcterization of a mammalian proton-coupled metal-ion transporter. Nature 1997; 388:482-488
9.
Garrick MD, Dolan KG, Horbinski C, Ghio AJ, Higgins D, Porubcin M, Moore EG, Hainsworth LN, Umbreit JN,
Conrad ME, Feng L, Lis A, Roth JA, Singleton S and Garrick LM. DMT1. A mammalian transporter for multiple
metals. BioMetals 2003; 16:41–54
10.
Hubert N, Hentze MW. Previously uncharacterized isoforms of divalent metal transporter (DMT)-1. implications
for regulation and cellular function. Proc Natl Acad Sci U S A. 2002; 99:12345-50
11.
McKie AT, Martinai P, Rolfs A, Brennan K, Wehr K, Barrow D, Miret S, Bomford A, Peters TJ, Farzaneh F,
Hediger MA, W. Hentze MW, Simpson RJ. A novel duodenal iron-regulated transporter, IREG1, implicated in the
basolateral transfer of iron to the circulation. Mol Cell 2000; 5:299-309
12.
Donovan A, Brownlie A, Zhou Y, Shepard J, Pratt SJ, Moynihan J, Paw BH, Drejer A, Barut B, Zapata A, Law
TC, Brugnara C, Lux SE, Pinkus GS, Pinkus JL, Kingsley PD, Palis J, Fleming MD, Andrews NC, Zon LI.
Positional cloning of zebrafish ferroportin1 identifies a conserved vertebrate iron exporter. Nature 2000;
403:776-781
13.
Vulpe CD, Kuo YM, Murphy TL, Cowley L, Askwith C, Libina N, Gitschier J, Anderson GJ. Hephaestin, a
ceruloplasmin homologue implicated in intestinal iron transport, is defective in the sla mouse. Nat Genet 1999;
21:195–199
14.
Conrad ME, and Umbreit JN. Iron absorption-The mucin mobilferrin integrin pathway. A competitive pathway for
iron absorption. Am J Hematol 1993; 42:67–73
15.
Hallberg L. Perspectives on nutritional iron deficiency. Annu Rev Nutr 2001; 21:1–21
16.
Worthington MT , Cohn SM, Miller SK, Luo RQ, Berg CL. Characterization of a human plasma membrane heme
transporter in intestinal and hepatocyte cell lines. Am J Physiol 2001; 280:G1172–G1177
17.
Weintraub LR, Conrad ME, Crosby WH. Absorption of hemoglobin iron by the rat. Proc Soc Exp Biol Med 1965;
120:840–843
18.
Boni RE, Huch Boni RA, Galbraith RA, Drummond GS, Kappas A. Tin-mesoporphyrin inhibits heme oxygenase
activity and heme-iron absorption in the intestine. Pharmacology 1993; 47:318–329
19.
Fleming RE, Sly WS. Mechanisms of iron accumulation in hereditary hemochromatosis. Annu Rev
Physiol.2002; 64:663-680
20.
Finch C. Regulators of iron balance in humans. Blood 1994; 84:1697-1702
21.
Krause A, Neitz S, Mägert HJ, Schulz A, Forssmann WG, Schulz-Knappe P, Adermann K. LEAP-1, a novel
highly disulfide-bonded human peptide, exhibits antimicrobial activity. FEBS Lett 2000; 480:147-150
22.
Pigeon C, Ilyin G, Courselaud B, Leroyer P, Turlin B, Brissot P, Loreal O. A new mouse liver-specific gene,
encoding a protein homologous to human antimicrobial peptide hepcidin, is overexpressed during iron overload.
J Biol Chem 2001; 276:7811–7819.
23.
Nicolas G, Bennoun M, Devaux I, Beaumont C, Grandchamp B, Kahn A, Vaulont S. Lack of hepcidin
gene expression and severe tissue iron overload in upstream stimulatory factor 2 (USF2) knockout mice. Proc
Natl Acad Sci USA 2001; 98:8780-8785
24.
Sandberg A-S, Brune M, Carlsson N-G, Hallberg L, Skoglund E, Rossander-Hulthén L. Inositol phosphates with
different numbers of phosphate groups influence iron absorption in humans. Am J Clin Nutr 1999; 70:240–246
25.
Kaltwasser JP, Werner E, Schalk K, Hansen C, Gottschalk R, Seidl C. Clinical trial on the effect of regular tea
drinking on iron accumulation in genetic haemochromatosis Gut 1998; 43:699-704
26.
Garcý´a-Casal MN, Leets I Layrisse M. ß-Carotene and Inhibitors of Iron Absorption Modify Iron Uptake by
Caco-2 Cells. J Nutr 2000; 130: 5–9
27.
Aisen P, Enns C, Wessling-Resnick M. Chemistry and biology of eukaryotic iron metabolism. Int J Biochem Cell
Biol. 2001; 33:940-959
28.
Ohgami RS, Campagna DR, Greer EL, Antiochos B, McDonald A, Jing Chen, Sharp JJ, Fujiwara Y, Barker JE,
Fleming MD. Identification of a ferrireductase required for efficient transferrin-dependent iron uptake in erythroid
cells. Nature Genetics 2007; 37:1264-69
29.
Kawabata H, Yang R, Hirama T, Vuong PT, Kawano S, Gombart AF, Koeffler HP. Molecular cloning of transferrin
receptor 2. A new member of the transferrin receptor-like family. J Biol Chem 1999; 274: 20826–20832
30.
Gao J, Chen J, Kramer M, Tsukamoto H, Zhang AS , Enns CA. Interaction of the Hereditary Hemochromatosis
Protein HFE with Transferrin Receptor 2 Is Required for Transferrin-Induced Hepcidin Expression. Cell Metab
2009; 9(3):217-27
31.
Mastroberardino PG, Hoffman EK, Horowitz MP, Betarbet R, Taylor G, Cheng D, Na HM, Gutekunst CA, Gearing
M, Trojanowski JQ, Anderson M, Chu CT, Peng J, Greenamyre JT. A novel transferrin/TfR2-mediated
mitochondrial iron transport system is disrupted in Parkinson's disease. Neurobiol Dis. 2009 Feb 26. [Epub
ahead of print
32.
Markovic M, Majkic-Singh N, Subota V. Usefulness of soluble transferrin receptor and ferritin in iron deficiency
and chronic disease. Scand J Clin Lab Invest. 2005; 65(7):571-576
33.
Brock JH. The physiology of lactoferrin. Biochem Cell Biol 2002; 80:1–6
34.
Suzuki YA and Lönnerdal B. Characterization of mammalian receptors for lactoferrin. Biochem Cell Biol 2002;
80:75-80
35.
Dunn LL, Sekyere EO, Rahmanto YS, Richardson DR . The function of melanotransferrin: a role in melanoma
cell proliferation and tumorigenesis. Carcinogenesis 7,2157–2169
36.
Fleming MD, Romano MA, Su MA, Garrick LM, Garrick MD, Andrews NC. Nramp2 is mutated in the anemic
Belgrade (b) rat: evidence of a role for Nramp2 in endosomal iron transport. Proc Natl Acad Sci USA 1998; 95:
1148–1153
37.
Roth JA, Horbinski C, Feng L, Dolan KG, Higgins D, Garrick MD. Differential localization of divalent metal
transporter 1 with and without iron response element in rat PC12 and sympathetic neuronal cells. J Neurosci
2000; 20:7595–7601
38.
Li JY, Paragas N, Ned RM, Qiu A, Viltard M, Leete T, Drexler IR, Chen X, Sanna-Cherchi S, Mohammed F,
Williams D, Lin CS, Schmidt-Ott KM, Andrews NC, Barasch J. Scara5 is a ferritin receptor mediating non-
transferrin iron delivery. Dev Cell. 2009; 16(1):35-46
39.
Fisher J, Devraj K, Ingram J, Slagle-Webb B, Madhankumar AB, Liu X, Klinger M, Simpson IA, Connor JR.
Ferritin: a novel mechanism for delivery of iron to the brain and other organs. Am J Physiol Cell Physiol 293:
C641–C649, 2007
40.
Todorich B, Zhang X, Slagle-Webb B, Seaman WE, Connor JR. Tim-2 is the receptor for H-ferritin on
oligodendrocytes. J Neurochem. 2008; 107(6):1495-50546.
41.
Oudit GY, Sun H, Trivieri MG, Koch SE, Dawood F, Ackerley C, Yazdanpanah M, Wilson GJ, Schwartz A, Liu PP,
Backx PH. L-type Ca2+ channels provide a major pathway for iron entry into cardiomyocytes in iron-overload
cardiomyopathy. Nature Medicine 9, 1187 - 1194 (2003)
42.
Gaasch JA, Geldenhuys WJ, Lockman PR, Allen DD, Van der Schyf VJ. Voltage-gated Calcium Channels
Provide an Alternate Route for Iron Uptake in Neuronal Cell Cultures. Neurochemical Research 2007; 32: 1686-
1693
43.
Ludwiczek, S., Theurl, I. , Muckenthaler, M.U., Jakab, M., Mair, S.M., Theurl, M., Kiss, J., Paulmichl, M.,
Hentze, M.W., Ritter, M. and Weiss, G. (2007) Ca2+ channel blockers reverse iron overload by a new
mechanism via divalent metal transporter-1. Nat. Med. 13, 448–454
44.
Napier I, Ponka P, and Richardson DR. Iron trafficking in the mitochondrion: novel pathways revealed by
disease. Blood. 2005 ;105:1867-1874
45.
Shaw GC, Cope JJ, Li L, Corson K, Hersey C, Ackermann GE, Gwynn B, Lambert AJ, Wingert RA, Traver D,
Trede NS, Barut BA, Zhou Y, Minet E, Donovan A, Brownlie A, Balzan R, Weiss MJ, Peters LL, Kaplan J, Zon LI,
Paw BH. Mitoferrin is essential for erythroid iron assimilation. Nature 2006; 440:96–100
46.
Levi S, Corsi B, Bosisio M, Invernizzi R, Volz A, Sanford D, Arosio P, Drysdale J. A human mitochondrial ferritin
encoded by an intronless gene. J Biol Chem 2001; 276: 24437-24440
47.
Nie G., Sheftel AD, Kim SF, and Ponka P. Overexpression of mitochondrial ferritin causes cytosolic iron
depletion and changes cellular iron homeostasis. Blood 2005; 5:2161-2167
48.
Santambrogio P, Biasiotto G, Sanvito F, Olivieri S, Arosio P, and Levi S. Mitochondrial ferritin expression in adult
mouse tissues. J Histochem Cytochem 2007; 55: 1129–1137.
49.
Bekri S, Kispal G, Lange H, Fitzsimons D, Tolmie J, Lill R, and Bishop DF. Human ABC7 transporter: Gene
structure and mutation causing X-linked sideroblastic anemia with ataxia with disruption of cytosolic iron-sulfur
protein maturation. Blood 2000; 96:3256-3264
50.
Campuzano V, Montermini L, Molto MD, Pianese L, Cossee M, Cavalcanti F, Monros E, Rodius F, Duclos F,
Monticelli A, Zara F, Canizares J, Koutnikova H, Bidichandani SI, Gellera C, Brice A, Trouillas P, De Michele G,
Filla A, De Frutos R, Palau F, Patel PI, Di Donato S, Mandel J-L, Cocozza S, Koenig M, Pandolfo M.
Friedreich's ataxia: autosomal recessive disease caused by an intronic GAA triplet repeat expansion. Science
1996; 271: 1423-1427
51.
Marmolino D, Acquaviva F. Friedreich's Ataxia: From the (GAA)( n ) Repeat Mediated Silencing to New
Promising Molecules for Therapy. Cerebellum. 2009 Jan 23 [Epub ahead of print].
52.
Millholland JM, Fitch JM, Cai CX, Gibney EP, Beazley KE, Linsenmayer TF. Ferritoid, a tissue-specific nuclear
transport protein for ferritin in corneal epithelial cells. J Biol Chem 2003; 278: 23963–23970
53.
Cai CX, Birk DE, and Linsenmayer TF. Nuclear ferritin protects DNA from UV damage in corneal epithelial cells.
Mol Biol Cell 1998; 9:1037–1051
54.
Harrison PM, Arosio P. The ferritins: molecular properties, iron storage function and cellular regulation. Biochim
Biophys Acta 1996; 1275:161-203
55.
Iancu TC. Ultrastructural pathology of iron overload. Bailliere's Clin Haematol 1989; 2: 475-49551.
56.
Muckenthaler MU, Galy B, Hentze MW. Systemic iron homeostasis and the iron-responsive element/iron-
regulatory protein (IRE/IRP) regulatory network. Annu Rev Nutr. 2008; 28:197-213.
57.
Kim S, Ponka P. 2000. Effects of interferon-gamma and lipopolysaccharide on macrophage iron metabolism are
mediated by nitric oxide-induced degradation of iron regulatory protein 2. J Biol Chem 2000; 275:6220–26
58.
Pantapoulos K. Iron metabolism and the IRE/IRP regulatory system. An Update. Ann NY Acad Sci 2004; 1012:1-
13
59.
Cooperman SS, Meyron-Holtz EG, Olivierre-Wilson H, Ghosh MC, McConnell JP, and Rouault TA. Microcytic
anemia, erythropoietic protoporphyria, and neurodegeneration in mice with targeted deletion of iron-regulatory
protein 2. Blood 2005; 106:1084-1091
60.
Finch C. Regulators of iron balance in humans. Blood 1994;84:1697–702.
61.
Deepak Darshan D, Anderson GJ. Interacting signals in the control of hepcidin expression. Biometals 2009;
22:77–87
62.
Theurl I, Aigner E, Theurl M, Nairz M, Seifert M, Schroll A, Sonnweber T, Eberwein L, Witcher DR, Murphy AT,
Wroblewski VJ, Wurz E, Datz C, Weiss G. Regulation of iron homeostasis in anemia of chronic disease and iron
deficiency anemia: diagnostic and therapeutic implications. Blood. 2009 Mar 17. [Epub ahead of print]
63.
Nemeth E, Tuttle MS, Powelson J, Vaughn MB, Donovan A, Ward DM, Ganz T, Kaplan J. Hepcidin regulates
cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004; 306: 2090–2093
64.
Domenicoa ID, Loa E, Diane M. Warda DM, Kaplana J. Hepcidin-induced internalization of ferroportin requires
binding and cooperative interaction with Jak2. PNAS 2009; 106: March 10 [Epub ahead of print]
65.
Swinkels DW, Girelli D, Laarakkers C, Kroot J, Campostrini N, Kemna EH, Tjalsma H. Improvements to
quantitative hepcidin measurements by mass spectrometry. PLoS One 2008;3: e2706.
66.
Ganz T, Olbina G, Girelli D, Nemeth E, Westerman M. Immunoassay for human serum hepcidin. Blood. 2008;
112:4292-4297
67.
Papanikolaou G, Samuels ME, Ludwig EH, MacDonald ML, Franchini PL, Dube MP, Andres L, MacFarlane J,
Sakellaropoulos N, Politou M, Nemeth E, Thompson J, Risler JK, Zaborowska C, Babakaiff R, Radomski CC,
Pape TD, Davidas O, Christakis J, Brissot P, Lockitch G, Ganz T, Hayden MR, Goldberg YP. Mutations in HFE2
cause iron overload in chromosome 1q-linked juvenile hemochromatosis. Nat Genet. 2004; 36:77-82
68.
Babitt JL, Huang FW, Wrighting DM, Xia Y, Sidis Y, Samad TA, Campagna JA, Chung RT, Schneyer AL, Woolf
CJ, Andrews NC, Lin HY. Bone morphogenetic protein signaling by hemojuvelin regulates hepcidin expression.
1: Nat Genet. 2006 May;38(5):531-9. Epub 2006
69.
Meynard D, Kautz L, Darnaud V, Canonne-Hergaux F, Coppin H, Roth M-P. Lack of the bone morphogenetic
protein BMP6 induces massive iron overload Nature Genetics 2009; 41, 478 – 481 | doi:10.1038/ng.320
70.
Andriopoulos B, Corradini E, Xia Y, Faasse SA, Chen S, Grgurevic L, Knutson MD, Pietrangelo A, Vukicevic S,
Lin HY, Babitt JL. BMP6 is a key endogenous regulator of hepcidin expression and iron metabolism. Nature
Genetics 2009; doi:10.1038/ng.335
71.
Du X, She E, Gelbart T, Truksa J, Lee P, Xia Y, Khovananth K, Mudd S, Mann N, Moresco EM, Beutler E,
Beutler B. The serine protease TMPRSS6 is required to sense iron deficiency. Science 2008;320:1088-92.
72.
Finberg KE, Heeney MM, Campagna DR, Aydinok Y, Pearson HA, Hartman KR, et al. Mutations in TMPRSS6
cause iron-refractory iron deficiency anemia (IRIDA). Nat Genet 2008;40:569-71
73.
Melis MA, Cau M, Congiu R, Sole G, Barella S, Cao A, Westerman M, Cazzola M, Galanello R. A mutation in the
TMPRSS6 gene, encoding a transmembrane serine protease that suppresses hepcidin production, in familial
iron deficiency anemia refractory to oral iron. Haematologica 2008; 93:1473-9











© www.eiseninfo.de
Eisenstoffwechsel
HepcidinHepcidin wurde zuerst als antimikrobiell wirksames Peptid im menschlichen Urin
isoliert und leap1 (Leber antimikrobielles Peptid) genannt (21). Bei einer knock-
out Maus wurde dann eher zufällig die Entwicklung einer
hämochromatosetypischen Eisenüberladung festgestellt und so ein
Zusammenhang zwischen dem Hamp-Gen (Hepcidin) und dem Eisenstoffwechsel
entdeckt (22,23). Inzwischen ist klar, dass Hepcidin den zentralen Regulator der
intestinalen Eisenabsorption darstellt (61, 62). Hepcidin bindet an IREG1 und
bewirkt eine Internalisierung dieses Eisenexporters, sodass der basolaterale
Eisentransport gedrosselt wird (63, 64). Die Synthese von Hepcidin in der Leber
ist abhängig von der Leber- und Plasmaeisenkonzentration, wird herunterreguliert
bei Eisenmangel und ist erhöht bei Eisenüberladung. Aber auch andere Faktoren
sind wichtig, die nicht unmittelbar etwas mit dem Eisenstoffwechsel zu tun haben
wie Sauerstoffmangel oder erhöhter Spiegel von Interleukin 6. Dies erklärt damit
auch die Hemmung der Eisenabsorption bei Hypoxie, Infektionen, Entzündungen
und Tumorerkrankungen. Hepcidin kann bisher nur mit relativ aufwendigen
Methoden im Serum und Urin von Patienten bestimmt werden, wird aber wohl
zukünftig an Bedeutung als neuer diagnostischer Parameter für eine Hemmung
der intestinalen Eisenabsorption gewinnen (65, 66).











